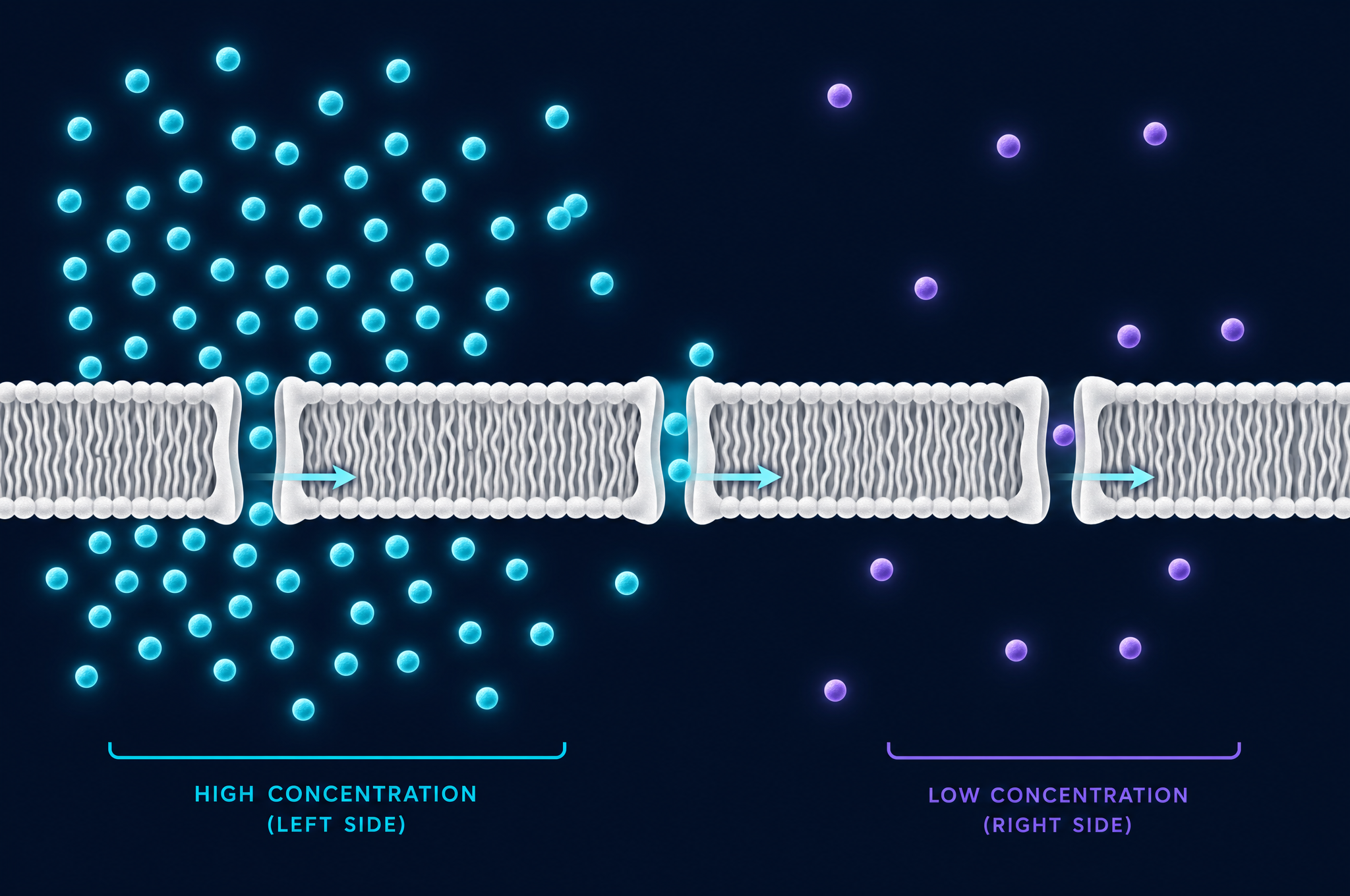
A neuron fires. A muscle contracts. A kidney cell filters your blood. Behind every one of these events is the same fundamental process: molecules moving across a membrane. The cell membrane is not a wall — it is a customs checkpoint, a gatekeeper, and a battery all in one. And the engine that drives most of the traffic across it is the same process that governs ink spreading through water: diffusion.
Understanding how cells manage molecular traffic is understanding how life maintains order in a universe that tends toward disorder. Every cell is a tiny pocket of organization — precise concentrations of ions, nutrients, and waste — existing in an environment that would dilute or flood it given the chance. The membrane is what holds that organization in place, and diffusion — both passive and actively managed — is what makes the exchange of materials possible.
The Membrane: A Lipid Bilayer
The cell membrane is built from a lipid bilayer — two layers of phospholipid molecules arranged tail-to-tail. Each phospholipid has a hydrophilic (water-loving) head and a hydrophobic (water-fearing) tail. In water, the tails naturally cluster together, forming a stable sheet with the heads facing outward toward the watery environments inside and outside the cell.
This bilayer is the key to the membrane's selectivity. Small, nonpolar molecules — like oxygen (O₂) and carbon dioxide (CO₂) — can slip directly through the lipid bilayer, because they are soluble in the hydrophobic interior. They cross by simple diffusion, moving from high concentration to low. This is how oxygen enters your cells and carbon dioxide leaves: no channels, no energy, just molecules diffusing down their concentration gradients through the lipid.
Oxygen is small and nonpolar, so it dissolves in the lipid interior of the membrane and diffuses through. Sodium (Na⁺) is an ion — charged and surrounded by a shell of water molecules. It cannot enter the hydrophobic lipid interior. To cross the membrane, ions need dedicated protein channels that provide a hydrophilic pathway through the lipid barrier.
Simple Diffusion: The Passive Pathway
For the molecules that can cross the lipid bilayer directly — oxygen, carbon dioxide, steroid hormones, some drugs — transport is by simple diffusion. The rate of transport is governed by Fick's first law: the flux is proportional to the concentration gradient, the membrane's permeability to the molecule, and inversely proportional to the membrane thickness.
This is why the alveolar membrane in your lungs is so thin — about 0.2 micrometers. The thinner the membrane, the faster oxygen can diffuse across it. Any thickening, whether from disease or scarring, slows oxygen transport and starves the tissues. The body's design is a direct application of Fick's law: minimize the diffusion distance to maximize the flux.
Simple diffusion is entirely passive — it requires no energy expenditure from the cell. The cell doesn't "do" anything to move oxygen in or carbon dioxide out. The gradients are maintained by the cell's metabolism (which consumes oxygen and produces CO₂) and by the circulatory system (which replenishes oxygen and removes CO₂). The cell simply lets diffusion do the work.
Facilitated Diffusion: Channels and Carriers
Most biologically important molecules — ions, sugars, amino acids — cannot cross the lipid bilayer directly because they are charged or too large. For these, the membrane provides protein pathways. This is facilitated diffusion: still passive (no energy required, movement is down the concentration gradient), but mediated by specific membrane proteins.
There are two main types:
- Channel proteins: Form pores through the membrane. Ions flow through these pores rapidly, driven by their electrochemical gradient. Aquaporins, which allow water to cross the membrane far faster than it could diffuse through the lipid, are channel proteins. So are the sodium and potassium channels that generate nerve impulses.
- Carrier proteins: Bind to specific molecules on one side of the membrane, undergo a conformational change, and release the molecule on the other side. The glucose transporter GLUT1 works this way, ferrying glucose into red blood cells. Carriers are slower than channels but more selective.
Facilitated diffusion shares an important property with simple diffusion: it is saturable. At low concentrations, the rate of transport increases with concentration (as Fick's law predicts). But at high concentrations, all the channels or carriers are occupied, and the rate plateaus — it cannot increase no matter how steep the gradient. This is the difference between diffusion through a lipid bilayer (which has no maximum rate) and diffusion through protein channels (which are limited by the number of available proteins).
The Electrochemical Gradient
For ions, diffusion isn't the only force at work. Ions are charged, so they are also influenced by the electrical potential across the membrane. The combination of the concentration gradient and the electrical gradient is called the electrochemical gradient. An ion will move in the direction that reduces both its concentration difference and its charge difference — and sometimes these two forces oppose each other.
Consider potassium (K⁺) in a typical neuron. The concentration inside the cell is about 30 times higher than outside, so the concentration gradient pushes potassium out. But the inside of the cell is negatively charged relative to the outside (about -70 mV), so the electrical gradient pulls the positively charged potassium back in. The net movement depends on the balance of these two forces — and this balance, called the equilibrium potential, is what determines the resting membrane voltage.
This is the same physics described by the Fokker-Planck equation: drift (the electrical force) and diffusion (the concentration gradient) combine to determine the net flux. The cell membrane is a living demonstration of drift-diffusion dynamics, with the added complexity that the cell actively maintains the gradients rather than letting them equilibrate.
Active Transport: Swimming Upstream
Here is the paradox: if diffusion always moves things down their concentration gradient, how does the cell maintain gradients that oppose diffusion? The sodium-potassium gradient, for instance, requires keeping sodium concentrations low inside the cell despite sodium constantly leaking in through channels. The answer is active transport — moving molecules against their gradient, which requires energy.
The most famous active transporter is the sodium-potassium pump (Na⁺/K⁺-ATPase). It uses energy from ATP — the cell's energy currency — to pump three sodium ions out of the cell while pulling two potassium ions in. This is the opposite of what diffusion would do: sodium is moved from low concentration (inside) to high concentration (outside), and potassium from high to low. The pump fights diffusion continuously, burning ATP to maintain the gradients that make nerve impulses, muscle contraction, and nutrient uptake possible.
Active transport is what distinguishes a living cell from a dead one. When a cell dies, ATP production ceases, the pumps stop, and the ion gradients gradually dissipate as everything diffuses to equilibrium. A dead cell's interior comes to match its exterior — no more sodium-potassium gradient, no more resting potential, no more excitability. Death is, in a sense, the victory of diffusion over active transport.
Osmosis: The Water Problem
Because the membrane is semipermeable — water crosses freely, but most solutes do not — the cell is subject to osmotic pressure. If the extracellular fluid becomes too dilute (hypotonic), water rushes in by osmosis and the cell swells. If it becomes too concentrated (hypertonic), water rushes out and the cell shrinks. Either extreme is dangerous.
Animal cells, which lack a rigid cell wall, are particularly vulnerable. This is why the body invests enormous energy in maintaining blood osmolarity within a narrow range. The kidneys, through a combination of filtration, active transport, and osmosis, constantly adjust the solute concentration of blood and extracellular fluid to keep cells in their isotonic comfort zone.
Endocytosis and Exocytosis: Bulk Transport
For very large molecules — proteins, entire particles — even facilitated diffusion is insufficient. These are moved by endocytosis (the membrane engulfs material, bringing it inside in a vesicle) and exocytosis (vesicles inside the cell fuse with the membrane, releasing their contents outside). These are active processes, requiring energy and molecular machinery, and they operate at a scale where individual diffusion is irrelevant.
Neurotransmitter release is a classic example. When a neuron fires, vesicles filled with neurotransmitter fuse with the cell membrane at the synapse, releasing their contents into the gap between neurons. The neurotransmitters then diffuse across the tiny synaptic cleft — about 20-40 nanometers — to bind receptors on the next neuron. Even at the scale of neural communication, diffusion is doing the final leg of the work.
The Membrane as a Diffusion Machine
What emerges from this catalog is a picture of the cell membrane as a sophisticated diffusion management system. It uses simple diffusion for small molecules that can cross the lipid. It uses facilitated diffusion for larger or charged molecules, providing selective protein pathways. It uses active transport to maintain the gradients that diffusion would otherwise erase. And it uses osmosis — a special case of diffusion — to manage water balance.
Every one of these processes ultimately traces back to the principles Adolf Fick described in 1855. The flux is proportional to the gradient. The rate depends on the diffusion coefficient, which depends on temperature, viscosity, and molecular size. The square-root-of-time scaling means diffusion works well over microscopic distances and fails over macroscopic ones — which is why cells are small and why multicellular organisms need circulatory systems.
The cell membrane is, in essence, a device for living with diffusion: letting it do the work where it can, supplementing it where it can't, and fighting it where it must. Life itself is a sustained negotiation with the second law of thermodynamics, and the membrane is where that negotiation happens, one diffusing molecule at a time.